西北高原所在 Molecular Biology and Evolution 发表论文揭示基因组结构变异在牛远缘杂交后代雄性不育中的潜在作用
雄性哺乳动物体内,精子是唯一能将亲本遗传和表观遗传信息传递给下一代的高度特化细胞。哺乳动物精子发生是一个高度复杂的细胞分化过程,涉及生殖细胞命运转变的动态调节,包括精原细胞的增殖和分化、精母细胞的减数分裂和精子变形三个主要阶段。相对于人和啮齿动物,家畜精子发生关键事件的分子调控不明确,涉及其中的细胞组成变化和基因表达动态大多未知。以牛为代表的家养动物为理解发生的保守性和差异性提供了独特的模型,绘制其生精细胞的分子图谱并剖析决定精子发生效率的关键因素,将有助于识别生育相关问题、发现新的雄性生育标志分子、开发提升繁殖效率的新策略,也将有助于鉴定导致普通牛和牦牛杂交后代雄性不育的因果基因。
中国科学院西北高原生物研究所动物繁育与功能基因组学团队通过组织形态学、减数分裂进程分析、单细胞转录组和基因组结构变异检测等实验,绘制了普通牛(Bos taurus)、牦牛(Bos grunniens)及其杂交后代犏牛和回交一代睾丸细胞各生精阶段生殖细胞及体细胞的转录特征,构建了发育轨迹以确定不同生精细胞的命运转变,同时也检测了牦牛和犏牛减数分裂中染色体参与的联会和同源重组进程,并筛选参与牛和牦牛杂种雄性不育的潜在调节因子。结果发现,普通牛和牦牛睾丸中有7种精原细胞,10种精母细胞和11种精子细胞亚型。相比之下,犏牛的睾丸中仅含有7种精原细胞亚型和6种初级精母细胞亚型。值得注意的是,犏牛精母细胞在第一次减数分裂前期的双线期到终变期转变时的停滞伴随着DNA双链断裂(Double-strand breaks, DSBs)修复的缺陷,且性小体在粗线期的形成受损。在回交一代的睾丸中,发现圆形精子和少量长性精子,表明生精阻滞现象得到显著缓解。单精子注射试验结果表明,回交一代睾丸中的长型精子显微授精后可支持胚胎发育。
研究团队通过联合分析揭示了一系列可能在调控牛精原细胞分化、减数分裂和精子生成中发挥重要作用的基因。通过比较分析,发现42.23%的基因在犏牛生精细胞差异表达但在回交一代中恢复,其中精母细胞中68.72%的差异表达基因得到恢复。通过检测X连锁基因在粗线期及双线期/次级精母细胞中的表达情况,结果发现,有40余个基因在犏牛中高表达,但是在回交后代中的表达水平与普通牛和牦牛相当,表明性染色体失活事件在犏牛粗线期精母细胞中发生异常,但在回交一代中得到恢复。这些发现表明,减数分裂I前期的缺陷是犏牛精子发生失败的主要原因,这些损伤很可能是由于生殖细胞中某些基因子集的失调所致。Dobzhansky-Muller模型预测,杂交不兼容源于至少两个在杂交物种中独立分化的基因之间的相互作用。因此,研究团队开展了转录组和基因组结构变异联合分析,发现在回交一代中恢复表达的661个基因携带有基因组结构变异,包括528个在犏牛生精细胞上调和133个下调基因。这些结构变异在精原细胞中与上调基因有关而在精母细胞中与下调基因有关。利用牛精母细胞蛋白组数据,研究团队鉴定了115个在犏牛精母细胞中表现出差异蛋白质丰度的基因。其中,24个携带基因组结构变异的基因在犏牛精母细胞中差异表达,但在回交后代的精母细胞中恢复,其中包括TDRD1,CLGN,CKD5RAP2和DDHD1等在精子发生中功能已知的基因。上述研究为进一步筛选影响犏牛精母细胞粗线期DSBs修复和性染色体失活的关键分子及其调控通路提供了数据集,从而在分子遗传学层面为犏牛雄性不育的诱因探寻和遗传矫正策略的制定奠定基础。同时,这项工作有助于阐明牛精子生成的分子调控网络,为大型哺乳动物的精子发生提供了重要的见解,并为探索大型动物物种形成和生殖隔离机制提供了重要信息。
相关研究结果以 Dynamic regulation of spermatogenesis and hybrid sterility revealed by single-cell analysis in yak and cattle 为题,于2026年2月2日在 Molecular Biology and Evolution(中国科学院1区TOP期刊)在线发表。西北高原所博士研究生伍仕鑫(已毕业)为论文第一作者,杨其恩研究员为通讯作者。该工作得到了国家重点研发计划(2021YFD1200405),国家自然科学基金(31972574 & U22A20447)和青海省自然科学基金团队(2025-ZJ-970T)项目的资助。
论文链接: https://doi.org/10.1093/molbev/msag027
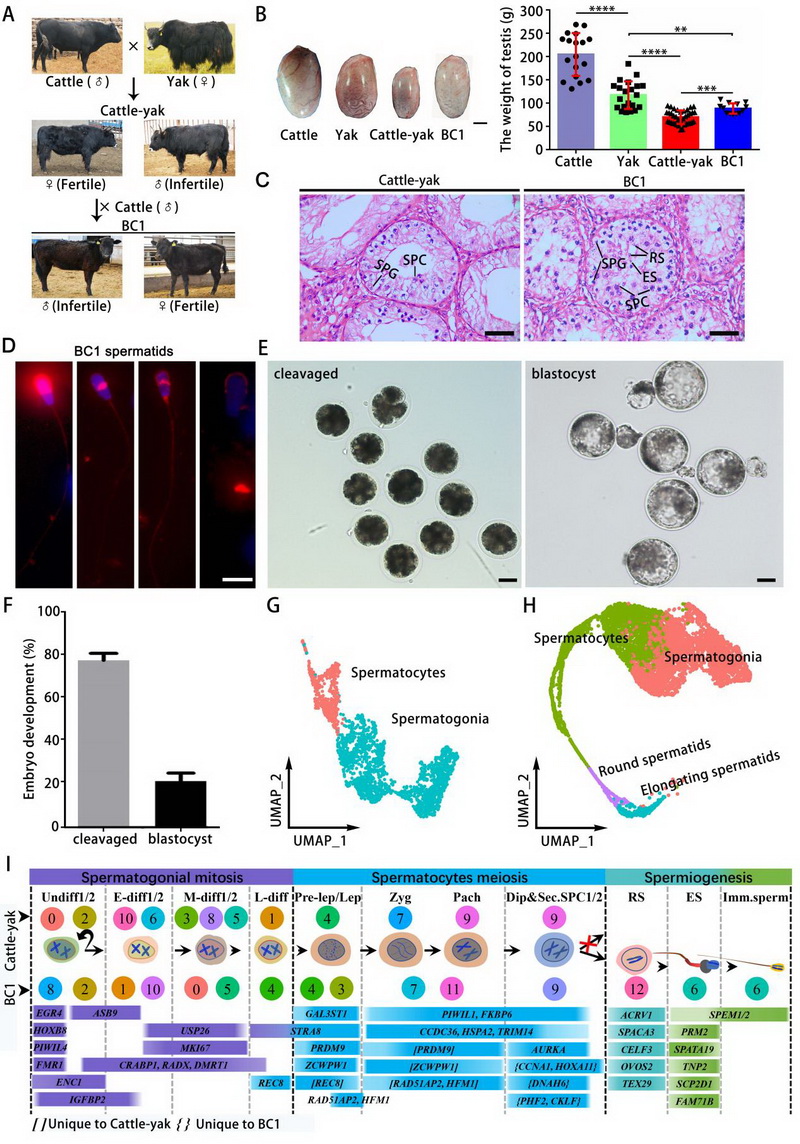
论文代表图1:普通牛和牦牛杂交后代和回交睾丸组织单细胞图谱分析。A) 普通牛与牦牛杂交产生犏牛及其回交后代(BC1)的模型示意图。B) 牦牛、家牛、犏牛及BC1的睾丸重量对比,比例尺 = 2 厘米;普通牛(n = 18)、牦牛(n = 21)、犏牛(n = 37)和BC1(n = 13)的睾丸重量进行两两比较。C) 犏牛(左)和BC1(右)睾丸组织的苏木精-伊红(H&E)染色切片。比例尺 = 50 μm。在犏牛中未观察到精母细胞阶段以后的生精细胞,而与犏牛相比,BC1的生精缺陷有所缓解。SPG:精原细胞,SPC:精母细胞,RS:圆形精子细胞,ES:伸长精子细胞。D) 从BC1睾丸中分离出的精子细胞顶体染色。比例尺 = 10 μm。E & F) BC1精子细胞胞质内注射入牦牛卵母细胞后获得了部分卵裂期胚胎和囊胚期胚胎;比例尺 = 50 μm。G) 犏牛生殖细胞的UMAP标注图,仅识别出两种亚型:精原细胞和精母细胞。H) BC1生殖细胞的UMAP标注图,识别出精原细胞、精母细胞、圆形精子细胞和伸长精子细胞。I) 犏牛和BC1生殖细胞各簇中特定标记基因的表达情况,可用于细胞类型鉴定。
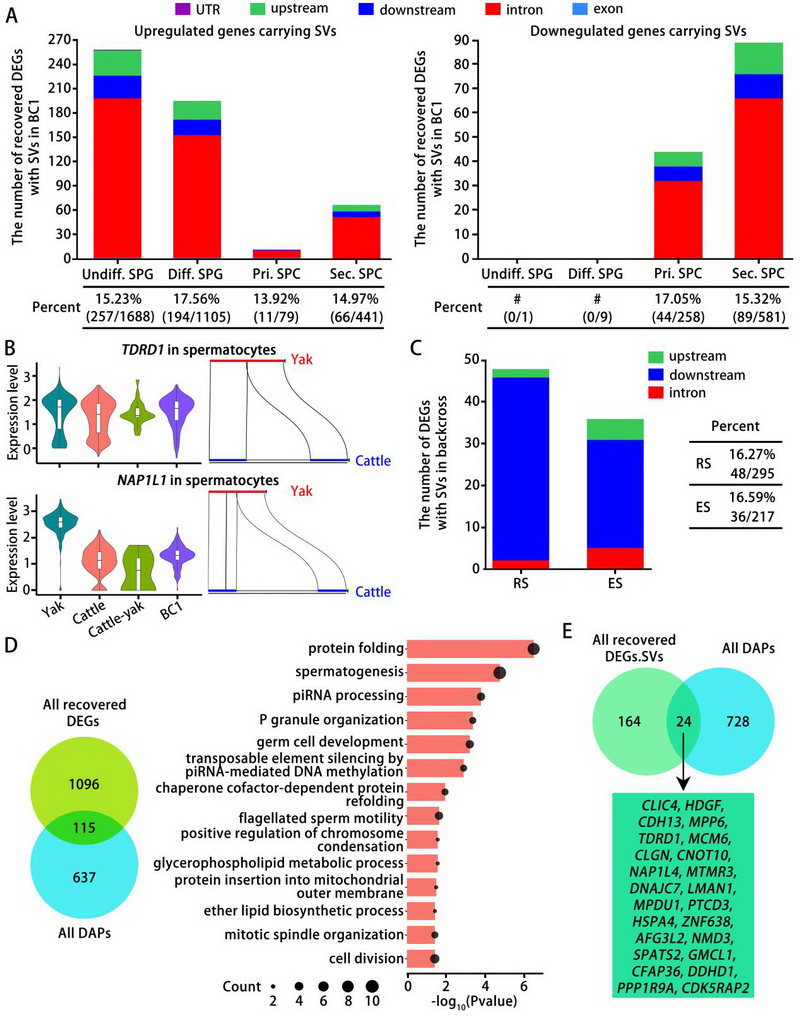
论文代表图2:普通牛、牦牛、犏牛及BC1各类生殖细胞亚型中携带基因组结构变异的差异表达基因。A) 柱状图显示在犏牛精原细胞和精母细胞各亚群中,具有结构变异且表达上调或下调、但在BC1中表达得以恢复的基因数量。B) 代表性基因的小提琴图及其结构变异的示意图。C) 柱状图显示与牦牛和普通牛相比,在BC1各类精子细胞亚型中携带基因组结构变异的差异表达基因数量。D) 对BC1精母细胞中所有表达水平恢复的差异表达基因,与犏牛精母细胞中的差异丰度蛋白进行韦恩图分析及GO富集分析。E) BC1精母细胞中所有表达恢复的差异表达基因、具有结构变异的差异表达基因,以及犏牛精母细胞中所有差异丰度蛋白的韦恩图分析。
 青公网安备 63010402000197号
青公网安备 63010402000197号